How Leishmaniasis Spreads Through Bites: Natural Sources Versus Lab Subjects
Jenn Hoskins
17th June, 2025



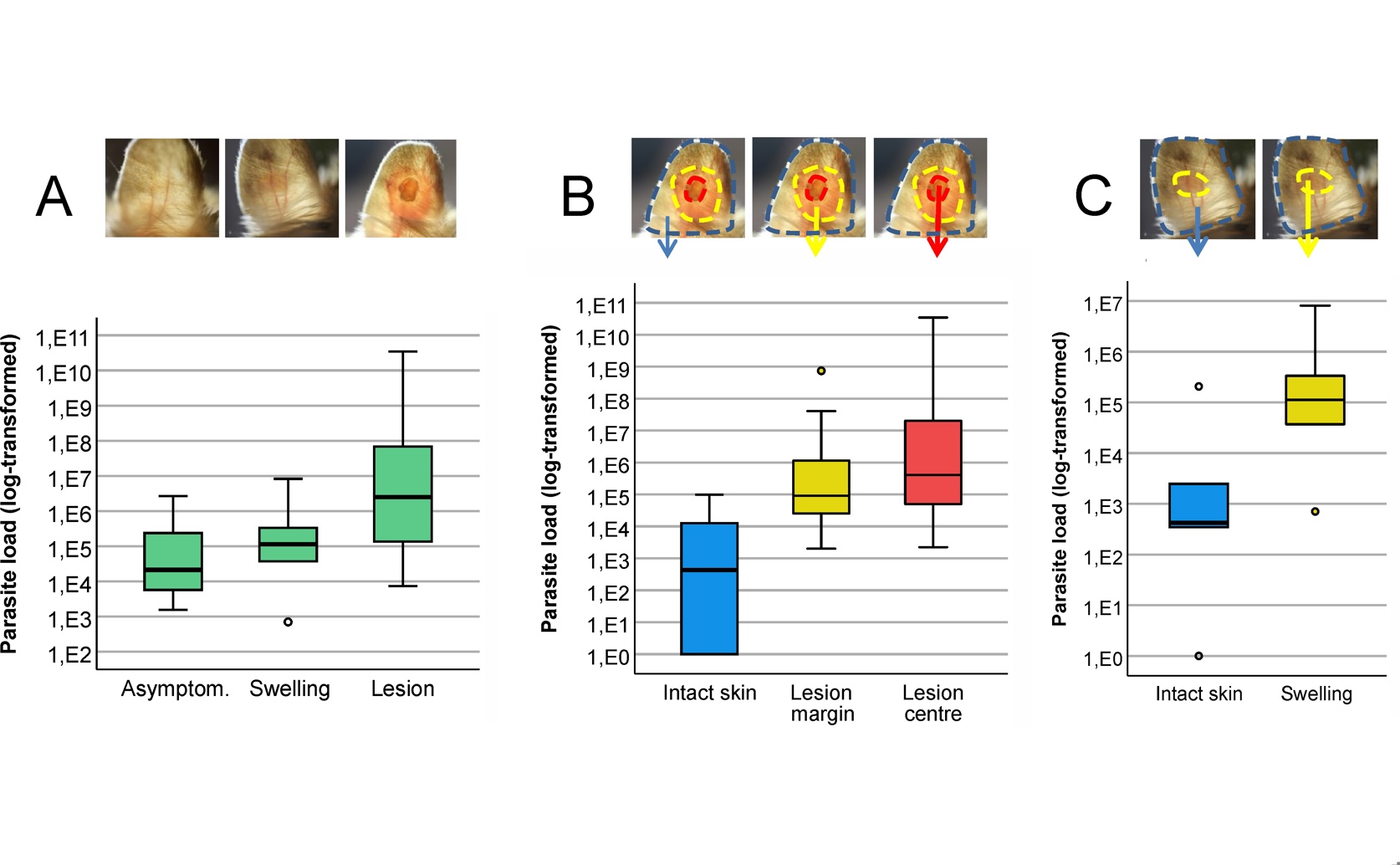
In its natural reservoir host, Shaw’s jird (Meriones shawi), Leishmania major parasites are heterogeneously distributed, concentrating in symptomatic areas of the ear (A) with parasite loads in lesion centers and margins being orders of magnitude higher than in the surrounding intact skin (B, C).
Key Findings
- In a study on leishmaniasis in North African rodents, parasites were clustered in skin lesions, with the edges being surprisingly more infectious to sand flies than the center
- Only a tiny number of parasites (2-10) were needed to infect sand flies from these natural hosts, a dose 100 times lower than required in lab mice
References
Main Study
1) Infectiousness of Leishmania major to Phlebotomus papatasi: differences between natural reservoir host Meriones shawi and laboratory model BALB/c mice
Published 16th June, 2025
https://doi.org/10.1371/journal.pntd.0013183
Related Studies
2) Skin parasite landscape determines host infectiousness in visceral leishmaniasis.
3) Infectiousness of Asymptomatic Meriones shawi, Reservoir Host of Leishmania major.
4) Minimally invasive microbiopsies: a novel sampling method for identifying asymptomatic, potentially infectious carriers of Leishmania donovani.
Related Articles
 1st June, 2025 | Jenn Hoskins
1st June, 2025 | Jenn Hoskins